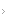 (Carregando Índice)... (Carregando Índice)... |
Autores:
Pedro Schestatsky
Médico Neurologista e Neurofisiologista. Especialista pela Academia Brasileira de Neurologia. Doutor em Ciências Médicas pela
UFRGS.
Artur Schuh
Médico Residente do Serviço de Neurologia do HCPA.
Josep Valls-Solé
MD/PHD. Consultor Sênior do Departamento de Neurologia do Instituto de Neurociências do Hospital Clinic, Barcelona.
Última revisão: 25/09/2014
Comentários de assinantes: 0
A eletroneuromiografia (ENMG) é o conjunto de exames complementares que permite examinar a função de várias estruturas do sistema nervoso central e periférico, devendo ser entendida como uma extensão do exame físico neurológico. De fato, existe uma significativa associação dos resultados da ENMG com o padrão de exame físico e o tipo de fibra nervosa lesionada (Lefaucheur; Créange, 2004), comprovando sua utilidade na prática clínica. A ENMG agrupa uma série de técnicas eletrodiagnósticas (Quadro 5.1) que o examinador deve ir selecionando e aplicando segundo seu próprio critério, com o intuito de fornecer informações sobre o estado global do sistema nervoso do paciente examinado. A exploração por ENMG deve ter como objetivo principal responder a questões levantadas durante a avaliação clínica do paciente pelo médico assistente. Para isso, é necessária a existência de um estreito contato entre o médico clínico que solicita o exame e o médico especializado que o realiza. Somente nessas condições a ENMG permitirá documentar e quantificar manifestações neurológicas que poderão ajudar no diagnóstico diferencial de certas síndromes neurológicas. De outra maneira, a exploração por ENMG pode gerar dados errôneos, potencialmente capazes de orientar tratamentos clínicos ou cirúrgicos desastrosos, como imunoglobulina endovenosa para polineuropatias, cirurgias desnecessárias para radiculopatias, canal estreito, compressões do nervo ulnar, etc.
As técnicas mais comumente utilizadas no eletrodiagnóstico convencional são a eletromiografia de agulha (EMG), a neurocondução motora e sensitiva, o estudo de reflexos segmentares e os potenciais evocados de curta latência. O eletromiografista dispõe também de técnicas relativamente sofisticadas para o estudo de determinados aspectos do controle motor, tais como tempo de reação (Nickerson, 1973; MacKay; Bonnet, 1990), reação de sobressalto (Brown et al., 1991; Valls-Solé et al., 1995), aprendizagem motora (Brasil-Neto et al., 1993) ou condicionamento clássico (WoodruffPak, 1988), que não serão abordadas no presente capítulo.
A máquina utilizada no eletrodiagnóstico se chama eletromiógrafo e possui um sistema de detecção e amplificação de sinais biológicos e um sistema de reprodução gráfica ou armazenamento. Um conjunto de estimuladores sincronizados com o tempo de análise do osciloscópio pode produzir estímulos elétricos em várias combinações ou ativar dispositivos externos complementares, dependendo da situação clínica particular.

Esse método consiste na avaliação funcional da atividade muscular por meio da inserção de um eletrodo de agulha na área do ponto motor de um determinado músculo. Dessa maneira, podem-se registrar potenciais de ação das fibras musculares despolarizadas pela chegada de impulsos de axônios motores. As agulhas comumente utilizadas são do tipo monopolar e captam a diferença de potencial entre a área central do bisel e a cânula, sob uma superfície relativamente ampla (150 x 580 micras). Em um músculo normal, o eletrodo de agulha não capta nenhuma atividade em repouso, com exceção de breves descargas de atividade irritativa no momento da inserção (atividade insercional fisiológica) e do possível achado de potenciais de placa motora, caso a agulha esteja situada em um terminal axonal. Durante a contração muscular, observam-se potenciais de ação unitários, denominados potenciais de unidade motora. Esses potenciais aumentam em frequência (somação temporal) e quantidade (somação espacial) ao aumentar a intensidade de contração. O conceito de unidade motora foi desenvolvido por Leyton e Sherrington, em 1925, para definir a unidade funcional integrada por um neurônio motor do corno anterior da medula, seu axônio e todas as fibras musculares inervadas por ele (Figura 5.1). A forma dos potenciais de unidade motora é variável: sua amplitude situa-se entre 0,5 e 2 mV, sua duração é de 6 a 10 ms, e seu número de fases, definido como o número de vezes em que a oscilação atravessa uma linha de base teórica, é de 2 a 4 vezes (Buchthal, 1957).
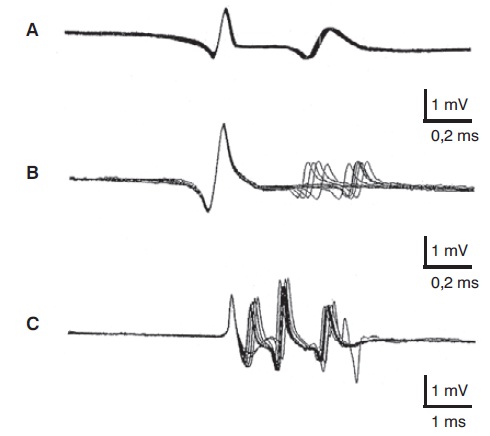
O recrutamento de um número cada vez maior de unidades motoras durante a contração muscular intensa dá lugar a um padrão de interferência registrado na tela do osciloscópio (Figura 5.2A). Por meio da técnica de turns/amplitude é possível analisar objetivamente esse padrão de interferência, contando-se o número de mudanças de direção oscilatória que apresentam uma amplitude superior a 100 µV e calculando-se sua amplitude média em um período de tempo pré-determinado (Daube, 1987). Em músculos de pacientes com neuropatia periférica,observam-se dois padrões fundamentais: aparecimento de atividade espontânea durante o repouso muscular e redução do número de potenciais de ação durante a contração muscular (Figura 5.2B). A atividade espontânea se manifesta por presença de potenciais de fibrilação, ondas agudas ou fasciculação. A redução do número de potenciais de unidade motora se traduz em uma menor densidade do padrão de interferência. Com frequência, os potenciais de ação mostram uma modificação na sua forma, aumentando o número de fases, a duração e a amplitude. Em músculos de pacientes com miopatia, pode-se observar também atividade espontânea em repouso na forma de fibrilação e ondas agudas, mas a mudança mais característica se observa durante a contração muscular. A perda funcional de fibras musculares da unidade motora dá lugar a uma deformação do potencial de ação, que mostra diminuição de sua amplitude, redução de sua duração e aumento do número de fases (Figura 5.2C).
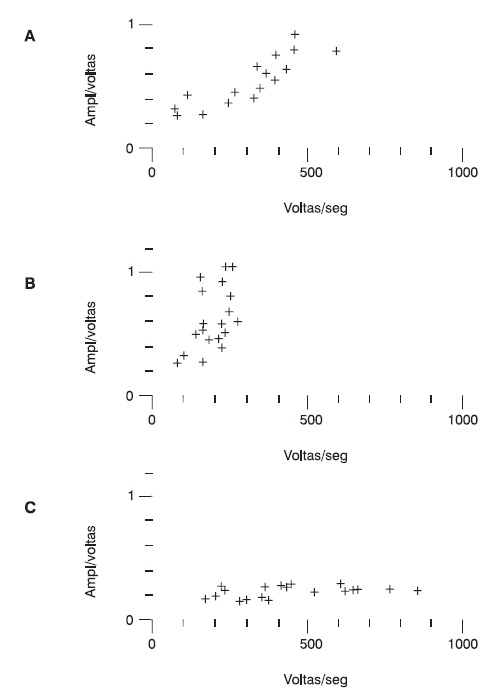
Outra consequência da perda funcional de fibras musculares é o recrutamento precoce do número de unidades motoras. Isso acarreta um padrão de interferência denso que já aparece com uma força escassa e um resultado no teste turns/amplitude bastante característico, utilizando o método de análise computadorizada (Figura 5.3).
Os eletrodos convencionais têm uma superfície maior do que o diâmetro médio das fibras musculares. Por isso, o potencial de ação registrado não permite identificar fibras musculares isoladas. Para esse objetivo, pode-se utilizar o eletrodo denominado fibra única (Stalberg; Trontelj, 1979), que possui uma superfície de registro de 25 micras de diâmetro. Após sua inserção no músculo do paciente, pede-se a ele que exerça uma contração muscular moderada e constante do músculo examinado. Devido à distribuição aleatória das fibras musculares de uma unidade motora, é registrado em 75% dos casos um potencial de ação único, de morfologia simples, trifásico e com duração total de aproximadamente 1 ms. Em 25% dos casos, o registro por meio de agulha de fibra única permite captar dois potenciais de ação sincronizados, que aparecem a certos intervalos de tempo entre eles (ativação de duas fibras musculares de uma mesma unidade motora) (Figura 5.4A). Em função da variabilidade da duração do potencial pós-sináptico excitatório que se gera na fibra muscular quando um impulso nervoso chega à placa motora, o tempo diferencial entre a ativação de uma fibra muscular e a de outra é variável. Essa variação se conhece como jitter e é de 24,6 ± 10,6 µs no músculo extensor comum dos dedos da mão (Stalberg; Trontelj, 1979). O jitter aumenta significativamente em doenças que cursam com diminuição do fator de segurança da transmissão neuromuscular, como a miastenia grave (Figura 5.4B). O jitter também aumenta em doenças neurogênicas ou miogênicas (Figura 5.4C), em que, além disso, pode-se observar aumento do número de potenciais de unidade motora durante a inserção da agulha, secundário à regeneração axonal e à reestruturação da unidade motora.
A atividade eletromiográfica também pode ser registrada com eletrodos de superfície. Esse tipo de registro é adequado para a análise da atividade muscular global ligada a movimentos voluntários ou involuntários (Dawson; Scott, 1949), tempo de reação (Nickerson, 1973; MacKay; Bonnet, 1990) ou reação de sobressalto (Brown et al., 1991; Valls-Solé et al., 1995).
Figura 5.1
Esquema visual da unidade motora, composta por: (1) corno anterior da medula, (2) raiz, (3) nervo espinal, (4) plexo, (5) nervo periférico, (6) junção neuromuscular e (7) fibras musculares (modificada de Dumitru, 1995).
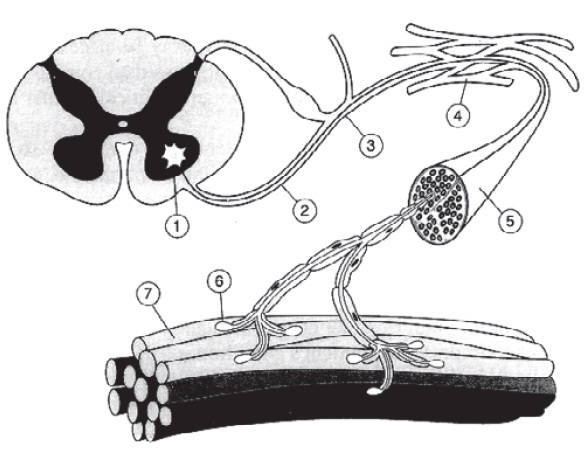
Figura 5.2
Eletromiografia do músculo normal (A), do músculo com desnervação por lesão neurogênica (B) e do músculo com afecção miogênica (C). Em cada caso, o traçado superior mostra o registro em repouso, e o inferior, o registro durante a contração muscular voluntária. Em repouso, observa-se silêncio elétrico no músculo normal e atividade espontânea no músculo desnervado (potenciais de fibrilação e ondas agudas) e no músculo com afecção miogênica (fibrilação). Durante a atividade muscular, observa-se um padrão interferencial no músculo saudável, um padrão deficitário no músculo desnervado e um padrão de interferência denso e de baixa amplitude no músculo com afecção miogênica.
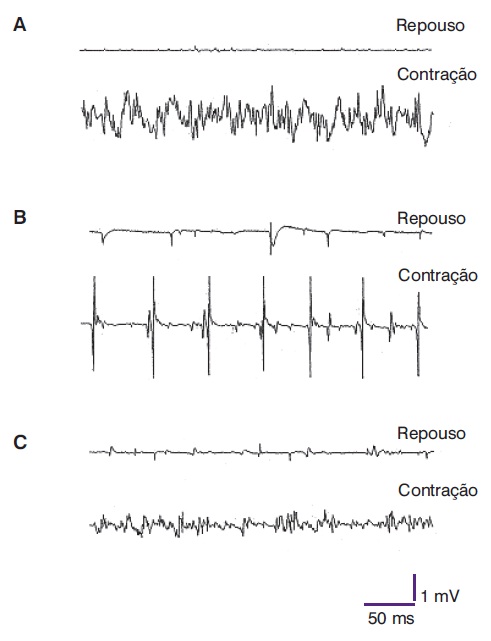
Figura 5.3
Representação da relação entre o número de voltas (eixo x) e a amplitude média das voltas (eixo y), obtida na análise computadorizada do padrão de interferência durante a contração voluntária em: (A) sujeito normal, (B) paciente com afecção neurogênica e (C) paciente com afecção miogênica.
Figura 5.4
Registro de EMG com agulha de fibra única. Todos os gráficos mostram uma superimposição de vários traçados contendo potenciais de ação da mesma unidade motora. (A) indivíduo normal, (B) paciente com miastenia grave e (C) paciente com desnervação crônica. Por esse método, o primeiro potencial de ação se mantém fixo na mesma posição, e os potenciais de ação da mesma unidade motora que se registram em seguida aparecem segundo sua relação temporal com o primeiro. Observe a maior variabilidade de representação do segundo potencial de ação em B com relação a A (aumento de jitter) e a maior quantidade de potenciais de ação em C em comparação com A e B (aumento de densidade de fibras secundário à reinervação compensatória).
A estimulação elétrica de um nervo motor produz a despolarização de seus axônios e gera impulsos ao longo do trajeto desse nervo. Quando esses impulsos alcançam o terminal axonal motor, e após a passagem pela junção neuromuscular, um potencial é propagado nas fibras musculares. Esse potencial pode ser registrado por meio de eletrodos de superfície ou de agulha, colocados sobre o músculo adequado. O estímulo elétrico utilizado para os estudos de neurocondução motora é, habitualmente, de intensidade supramáxima, ou seja, um estímulo capaz de despolarizar a maioria dos axônios de um nervo, ativando sincronizadamente a maioria das unidades motoras. O potencial resultante é denominado potencial evocado motor composto (PEMC) ou onda M. Quando se aplicam dois estímulos em pontos distintos de um mesmo nervo e se registra a onda M em um ponto distal, pode-se calcular a velocidade de condução das fibras motoras do nervo em questão entre os dois pontos de estimulação, dividindo-se a distância entre eles (em mm) pela latência diferencial (em ms) entre as respostas obtidas. A velocidade de condução normal é de aproximadamente 50 m/s. Frente à suspeita de uma doença da junção neuromuscular, estímulos elétricos repetidos a uma frequência de 3 Hz são efetuados sobre o nervo acessório com registro de PEMC no músculo trapézio. A observação de um decremento de amplitude superior a 10% dos potenciais de ação subsequentes com relação ao primeiro da série é altamente sugestiva de miastenia grave ou de síndrome de Lambert-Eaton (Scherer; Bedlack; Simel, 2005).
Para a neurocondução sensitiva, o estímulo se aplica sobre nervos sensitivos, com regis
tro por meio de eletrodos de superfície localizados no trajeto do nervo em questão. A neurocondução mista consiste em estimulação de um nervo misto e registro da resposta em um outro ponto do mesmo nervo. O potencial de ação registrado sobre o trajeto do nervo (denominado potencial de ação do nervo sensitivo, ou PANS) é de amplitude muito menor do que se registra no músculo (PEMC). Consequentemente, para melhorar as condições de registro da neurocondução sensitiva ou mista, emprega-se a técnica de promediação elétrica contínua de sinais consecutivos. Desenvolvido por Dawson e Scott (1949), esse sistema permite o cancelamento de oscilações surgidas ao acaso e produz, conseqüentemente, o aumento da nitidez da resposta final.
O estudo das neuroconduções sensitiva e motora é parte fundamental no diagnóstico das polineuropatias.
A despolarização de um axônio por meio de estímulo elétrico dá lugar a um potencial de ação propagado em duas direções do axônio (ortodrômica e antidrômica). Os impulsos conduzidos em fibras motoras e sensitivas em direção distal-proximal podem desencadear respostas musculares por ativação dos próprios neurônios motores. Essa ativação pode ser gerada por via transináptica, a partir de impulsos excitatórios que chegam ao corpo do neurônio motor por meio de axônios sensitivos (onda H), ou por via antidrômica, a partir da invasão do corpo do neurônio motor por impulsos propagados nos próprios neurônios motores (onda F). A onda H é um verdadeiro reflexo sensitivo motor, enquanto a onda F não depende de integridade da via sensitiva para sua produção. A importância clínica do estudo das respostas H e F se dá pela possibilidade de avaliar a condução em segmentos mais proximais do nervo, incluindo a porção intraraquídea da raiz nervosa (porção pré-ganglionar, especialmente comprometida nas radiculopatias).
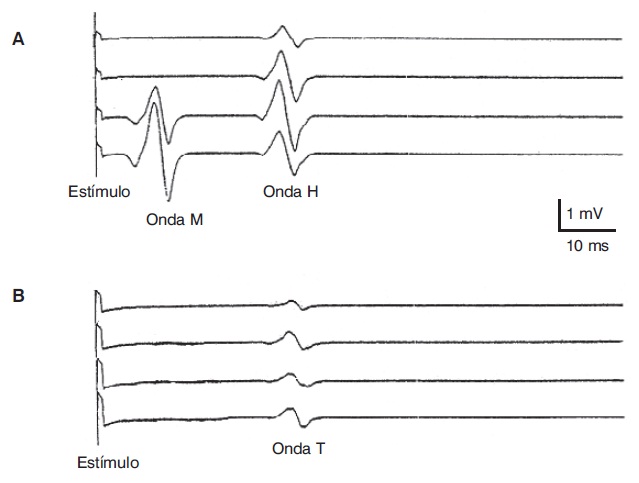
A resposta H pode ser obtida somente em alguns músculos do corpo humano: sóleo e quadríceps nas extremidades inferiores e flexor comum dos dedos nas extremidades superiores. A onda H é o equivalente eletrofisiológico do reflexo monossináptico de estiramento. O potencial de ação induzido pela percussão tendínea pode ser examinado caso seja utilizado um martelo de reflexos que seja capaz de disparar o varrido do osciloscópio do eletromiógrafo no momento da percussão. A onda gerada dessa forma se chama onda T (Figura 5.5). Ainda que as ondas H e T tenham trajetos aferente e eferente comuns, existem diferenças bem definidas entre elas, devido ao fato de que a onda T gera um impulso aferente de modo fisiológico, por ativação dos receptores de estiramento, enquanto o estímulo elétrico produz uma ativação direta do axônio (Schieppati, 1987). Por esse motivo, uma ausência de onda T com preservação de onda H sugere neuropatia distal incipiente, já que os receptores sensitivos são os primeiros a serem acometidos nessas situações.
A onda F pode ser obtida na maioria dos músculos distais das extremidades. A latência das respostas H e F está diretamente relacionada a medidas antropométricas, tais como a longitude do segmento estudado e o tamanho do sujeito. Existem fórmulas para medir a velocidade de condução de segmentos nervosos proximais. No caso da onda F, a velocidade de condução pode ser medida utilizando-se a seguinte fórmula:
Distância vértebra-estímulo × 2 (mm) / latência F – M – 1 (ms)
Nessa fórmula, a distância vértebra-estímulo se mede entre a vértebra correspondente (C6 para extremidades superiores e L1 para extremidades inferiores) e o ponto de estímulo (punho para as extremidades superiores e tornozelo para as extremidades inferiores). Essa distância deve ser multiplicada por dois, já que os impulsos correm em ambas as direções. A latência diferencial entre F e M deve ser reduzida em 1 ms, tempo estimado para que impulsos antidrômicos possam reexcitar os neurônios motores.
Com exceção de algumas provas que envolvem os nervos facial e acessório, os nervos cranianos não são acessíveis ao estudo eletrofisiológico por meio de métodos de neurocondução convencionais. Em função disso, a maior parte de informação sobre os nervos cranianos provém de estudos de respostas reflexas. Os reflexos de tronco cerebral dividem-se em reflexos trigêmino-trigeminais e reflexos trigeminofaciais. A maioria é polissináptica e envolve atividade de interneurônios em cadeia. Visto que a excitabilidade dos interneurônios do tronco cerebral é regulada por estruturas de hierarquia superior no sistema nervoso central, o estudo neurofisiológico dos reflexos de tronco contribui com informação útil sobre a excitabilidade interneuronal e doenças neurodegenerativas (Cruccu et al., 2005).
O reflexo mandibular é o único reflexo monossináptico de nervos cranianos acessível à análise neurofisiológica. É obtido por meio da percussão da mandíbula mediante um martelo de reflexos. Diferentemente de outros reflexos osteotendinosos, a exploração clínica do reflexo mandibular não permite ao investigador distinguir uma resposta normal de uma outra diminuída ou ausente. Entretanto, o registro eletrofisiológico, por meio de eletrodos colocados sobre o ponto motor do músculo masseter ao nível do ângulo da mandíbula, permite efetuar uma avaliação objetiva da resposta (Ongerboer de Visser; Goor, 1976). Uma propriedade intrínseca do reflexo mandibular é que o corpo neuronal dos axônios que veiculam a informação gerada nos receptores de fusos musculares encontra-se no núcleo mesencefálico, dentro do tronco cerebral (intra-axial). Esse é um aspecto diferencial da fisiologia do nervo trigêmeo, distinto de qualquer outro nervo do corpo humano, e permite explicar certos sinais clínicos de algumas doenças neurológicas (Valls-Solé et al., 1990).
Durante a contração sustentada, estímulos ao redor da boca, dos dentes ou da mucosa gengival produzem uma supressão transitória da atividade eletromiográfica, fenômeno denominado de reflexo massetérico inibitório (RMI). O estímulo mais utilizado na prática para induzir o RMI é o estímulo elétrico do nervo mentual (Ongerboer de Visser et al., 1990).
Figura 5.5
Ondas H e T do músculo sóleo no indivíduo normal.
(A) A onda H aparece com baixa intensidade de estímulo e diminui ao aumentar sua intensidade, quando surge a onda M. (B) A onda T aparece depois da percussão mecânica com um martelo de reflexos sincronizados com o varrido do osciloscópio, e sua latência é levemente maior do que a onda H.
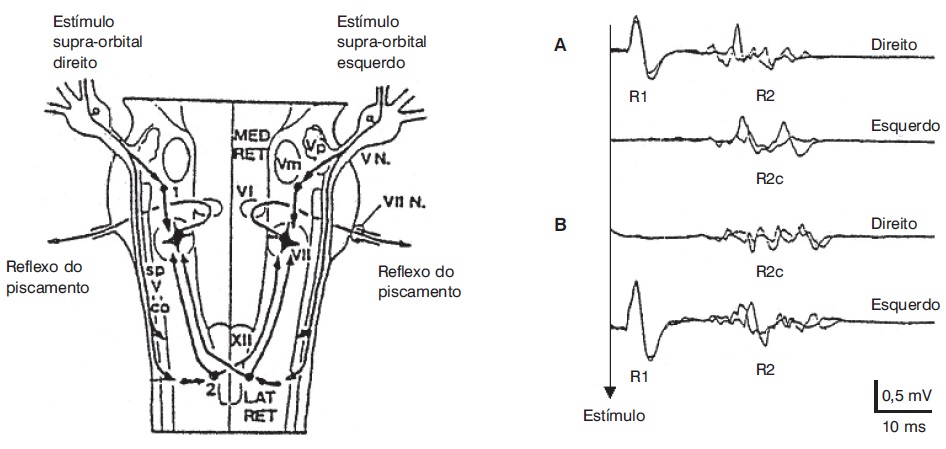
O estímulo provocado à córnea, por exemplo, por contato de uma gase ou um jato de ar, provoca o fechamento palpebral. Outros estímulos periorbitários provocam também a mesma resposta. O reflexo do piscamento, ou blink reflex, é uma ferramenta útil no estudo neurofisiológico, não somente das vias envolvidas no seu próprio circuito (Kimura; Powers; Van Allen, 1969; Shahani, 1970), mas também da excitabilidade neuronal suprasegmentar (Kimura, 1973; Valls-Solé; Lou; Hallett, 1994). O método mais comumente utilizado é o estímulo elétrico unilateral do nervo supraorbital com registro de respostas por meio de eletrodos de superfície colocados sobre o músculo orbicular dos olhos. Por meio dessa montagem, pode-se obter uma resposta ipsilateral, formada por dois componentes (R1 e R2), e uma resposta contralateral, formada por um só componente (R2c). A Figura 5.6 mostra o circuito de cada uma dessas respostas através do tronco cerebral. A estimulação digital da zona perioral produz um reflexo de protrusão labial, que também é acessível ao estudo eletrofisiológico (reflexo perioral). Para a sua obtenção, pode-se aplicar um estímulo mecânico ou elétrico no lábio superior e registrá-lo com eletrodos de superfície colocados na comissura labial (Topka; Hallett, 1992).
Figura 5.6
Reflexo do piscamento em um indivíduo saudável. À esquerda: esquema de circuitos no tronco cerebral pelas aferências trigeminais. À direita: respostas obtidas por estímulo do nervo supraorbital direito (A) e esquerdo (B) em um indivíduo saudável.
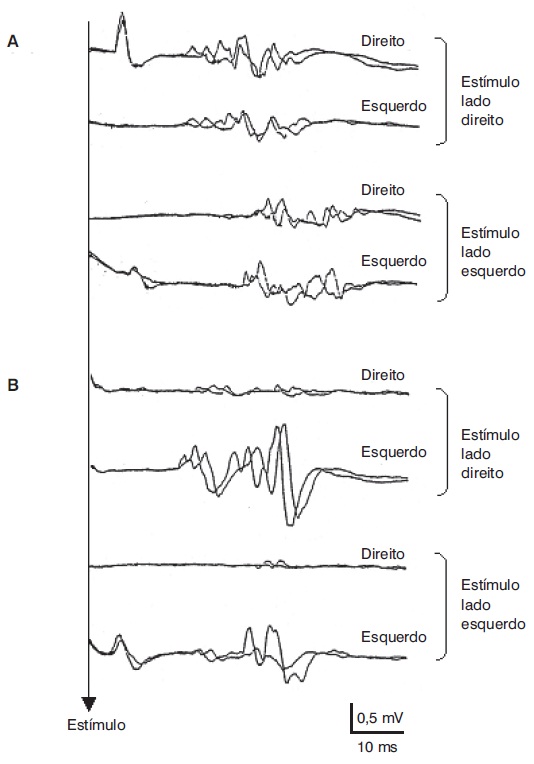
O reflexo do piscamento e o RMI são consensuais, ou seja, o estímulo do nervo de um lado da face produz respostas em músculos dos dois lados. Em função disso, essas respostas são muito úteis para reconhecer afecções dos aferentes sensitivos ou eferentes motores no segmente correspondente (primeiro ramo do trigêmeo e nervo facial para o reflexo do piscamento e terceiro ramo do trigêmeo e ramo motor do trigêmeo para o RMI). Nos casos de lesão da via aferente, o estímulo do lado afetado não dará lugar a respostas em nenhum dos lados, e o estímulo no lado saudável produzirá respostas normais (padrão aferente). Nos casos de lesão na via eferente, o estímulo aplicado em qualquer lado produzirá respostas normais no lado saudável e nenhuma resposta no lado afetado (Figura 5.7).
O estudo eletrofisiológico do sistema nervoso autonômico pode implicar o uso de métodos bastante complexos. No presente capítulo, nos limitaremos a descrever os testes mais utilizados nos laboratórios de neurofisiologia: a resposta simpáticocutânea (RSC) e a variação do intervalo R-R da frequência cardíaca.
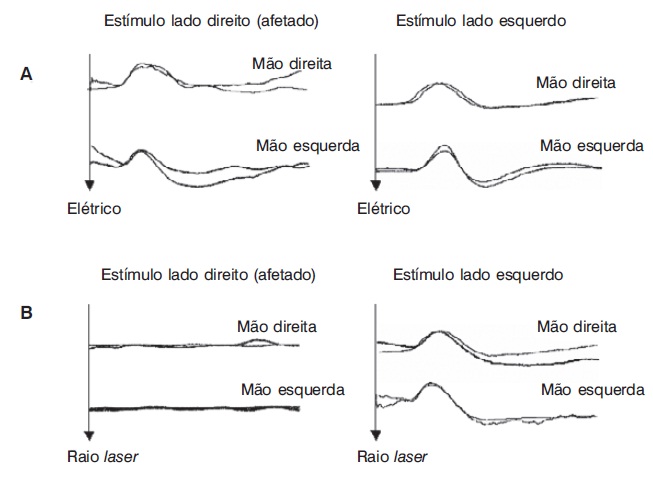
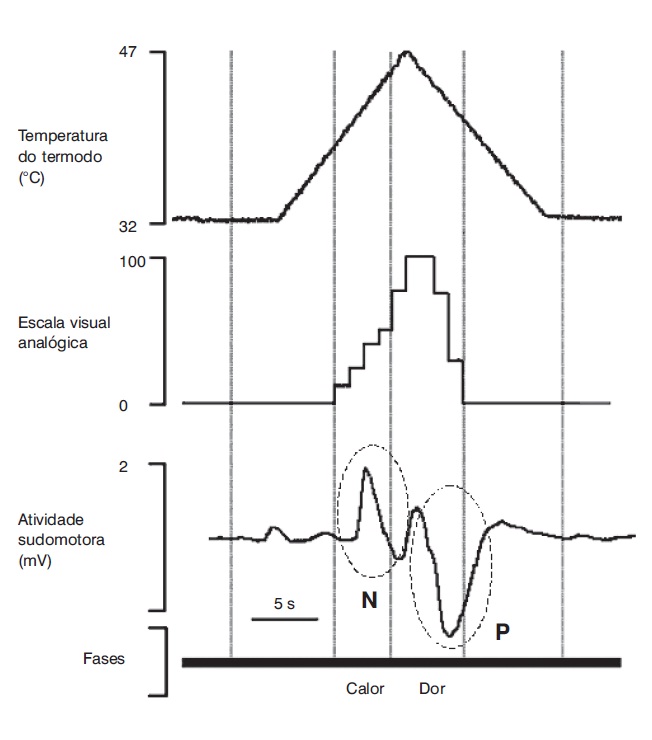
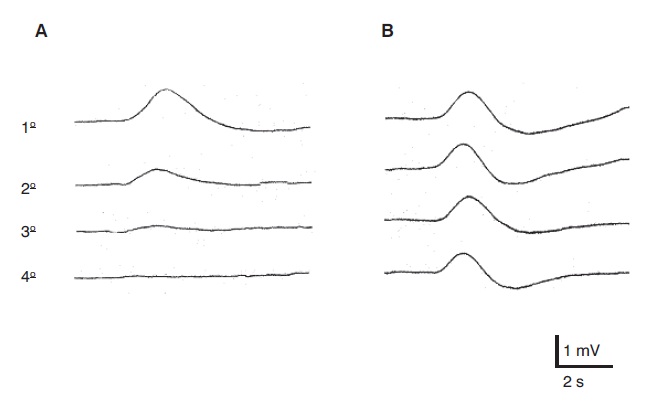
O estudo da RSC pode indicar o estado funcional das fibras C simpáticas eferentes. Eletrodos de registro são colocados na palma (ativo) e no dorso da mão (referência), e estímulos de modalidades diversas podem ser aplicados. O estímulo mais utilizado na prática é o elétrico, sobre o nervo periférico. Com ele se obtém uma resposta de lento desenvolvimento, a uma latência de aproximada mente 1,5 s, resultante de uma mudança na resistência galvânica cutânea sob o eletrodo de registro, produzida por uma ativação sincronizada de glândulas sudoríferas. Utilizando-se diferentes pontos de estímulo e de registro simultâneo da RSC, é possível determinar a localização da lesão na via aferente ou eferente do reflexo sudomotor (Montagna; Liguori; Zappia, 1985; Valls-Solé; Monforte; Estruch, 1991). A Figura 5.8 mostra RSCs evocadas por estímulo elétrico (A) e por raio laser (B), em um paciente com siringomielia (Veciana et al., 2007), demonstrando preservação da coluna dorsal e lesão seletiva da via espinotalâmica direita. Mais recentemente, a análise da morfologia dos potenciais tem sido proposta para auxiliar na interpretação de testes psicofísicos que avaliam a percepção de dor e temperatura (Figura 5.9) em sujeitos normais (Schestatsky et al., 2007a) e seu padrão de habituação (Figura 5.10) para avaliação funcional da via descendente inibidora da dor em pacientes com dor neuropática do tipo central (Schestatsky et al., 2007b).
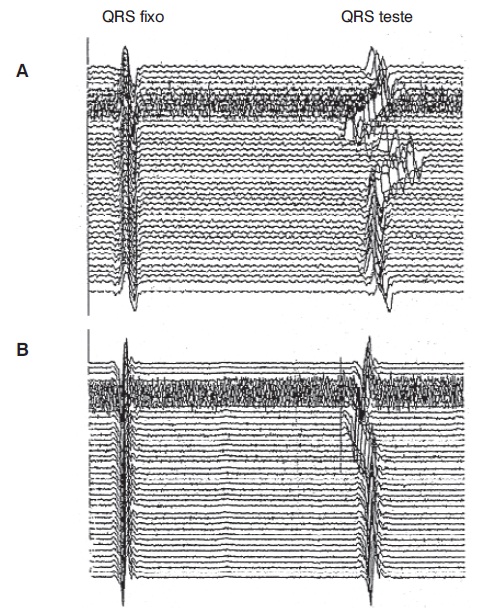
O estudo da variação do intervalo R-R em sucessivas batidas cardíacas pode indicar o estado funcional de fibras do sistema parassimpático. O complexo QRS do eletrocardiograma é utilizado para sincronizar o traçado do osciloscópio, de maneira que a posição que representa o próximo complexo QRS mostrará uma variabilidade diretamente relacionada com a variabilidade fisiológica da frequência cardíaca (Figura 5.11). Diversas manobras permitem efetuar um estudo funcional da variação do intervalo R-R, tais como a respiração profunda, a manobra de Valsalva ou a passagem do decúbito ao ortostatismo. Em pacientes cuja condição física não permite uma boa colaboração, é possível examinar a variabilidade do intervalo R-R durante a respiração tranquila em decúbito (Navarro, 1992).
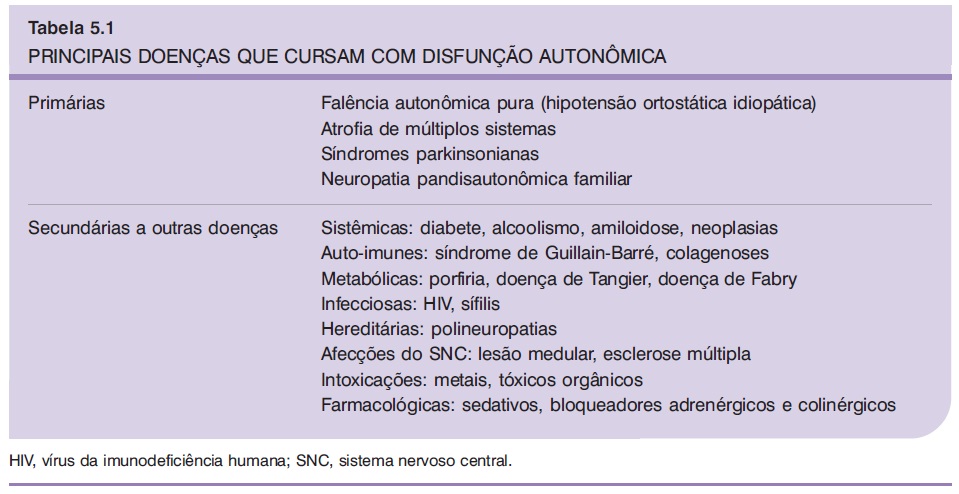
A Tabela 5.1 mostra uma lista de síndromes que cursam com disfunção do sistema nervoso autonômico em que um estudo neurofisiológico pode ser útil (Navarro, 1992).
Figura 5.7
(A) Reflexo do piscamento obtido por estímulo do nervo supraorbital em paciente com lesão trigeminal esquerda (padrão aferente). (B) Lesão do nervo facial direito (padrão eferente). Note respostas de maior latência quando o estímulo é aplicado no lado esquerdo (A) e ausência de qualquer resposta no lado direito por estímulo em qualquer dos lados (B).
O estímulo sensorial evoca uma atividade cerebral que pode ser detectada utilizando-se técnicas adequadas. Conforme o tipo de estímulo empregado, os potenciais evocados podem ser visuais, auditivos, somatossensoriais (todos esses induzidos por estímulo elétrico) ou nociceptivos (induzidos por temperatura ou raio laser). O Capítulo 6 fornece informações mais detalhadas sobre esses métodos.
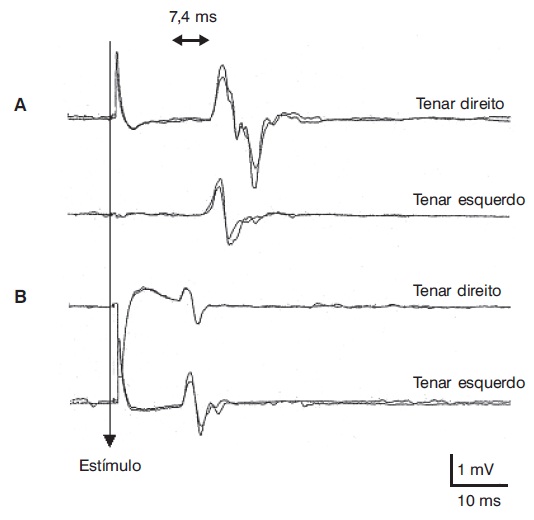
A estimulação do córtex cerebral foi incorporada recentemente ao conjunto de métodos de estudo eletrofisiológico (Chokroverty, 1989). O estímulo elétrico de alta intensidade cedeu lugar ao estímulo magnético transcraniano (EMT), que, por sua vez, é capaz de ativar regiões cerebrais de maneira não invasiva e não dolorosa (Barker; Jalinous; Freeston, 1985; Rothwell et al., 1991). Esse método se baseia na descarga sincronizada de energia armazenada em um banco de capacitores através de uma bobina de fio de cobre, que se posiciona sobre áreas específicas da cabeça do sujeito examinado. Quando se aplica a bobina sobre a região correspondente do córtex motor, obtém-se a geração de uma resposta muscular em vários músculos do organismo. Também pode-se gerar uma resposta nos mesmos músculos por aplicação de estímulo no nível da coluna vertebral e consequente despolarização de células do trato corticospinal e de raízes nervosas motoras dessa área no nível medular correspondente. A diferença de latência da resposta gerada por ativação cortical e medular se conhece como tempo de condução eferente central, que, em indivíduos normais em repouso, supera 9,5 ms nos músculos da mão e 18 ms nos da perna (Figura 5.12).
O estímulo cortical dá lugar a potenciais evocados motores (PEM), que se geram depois da despolarização da via corticospinal e da conseguinte excitação do neurônio motor espinal por conexões diretas (via córtico-motoneuronal) ou mediadas por interneurônios espinais. Assim, a resposta gerada por estímulo cortical depende não somente da excitabilidade do córtex motor, mas também dos neurônios motores espinais. O EMT sozinho, repetitivo ou combinado com outros métodos de ativação do neurônio motor fornece uma informação de grande importância para o estudo da fisiologia motora no ser humano, além de oferecer opção terapêutica em casos selecionados de dor neuropática, transtornos de movimento ou depressão maior intratáveis (Kobayashi; Pascual-Leone, 2003).
Figura 5.8
Respostas simpático-cutâneas induzidas por estímulo elétrico (A) e por raio laser (B) em paciente com siringomielia e dor neuropática. Observe respostas normais em (A), por estímulos em ambos os lados, e ausentes em (B), quando o estímulo é do lado direito. Esse padrão indica preservação da coluna dorsal bilateral e lesão no sistema ânterolateral direito (lesão aferente do trato espinotalâmico) (Fonte: Veciana et al., 2007).
Figura 5.9
Registro simultâneo do aumento da temperatura de um termodo colocado sobre a pele do antebraço, da percepção subjetiva dessa temperatura e da atividade eletrodérmica cutânea em um sujeito normal. Observe a presença de um potencial sudomotor negativo (onda N) no momento de percepção de calor e de um potencial sudomotor positivo (onda P) relacionado à percepção de dor por calor (Fonte: Schestatsky et al., 2007a).
Figura 5.10
Padrão de habituação de resposta sudomotora induzida por quatro estímulos repetidos de raio laser em sujeitos normais (A) e em pacientes com dor neuropática do tipo central (B) (modificado de Schestatsky et al., 2007b). Observe a falta de habituação nos pacientes com dor neuropática.
Figura 5.11
Registro da variação da frequência cardíaca durante a realização da manobra de Valsalva em sujeito normal (A) e em paciente com disfunção autonômica grave associada a polineuropatia diabética (B). Observe a menor variação do intervalo R-R em (B).
Figura 5.12
Respostas musculares evocadas por estímulo magnético cortical (A) e medular cervical (B), registradas em musculatura tenar de um sujeito saudável. O tempo de condução central calculado (valor da latência cortical menos a latência cervical medular) é de 7,4 ms (normal até 9,5 ms).
A eletroneuromiografia é um método diagnóstico útil, barato e não invasivo na avaliação funcional de doenças neurológicas do sistema nervoso periférico e/ou central. Tal técnica, além do auxílio diagnóstico, permite ao neurologista melhor entendimento da fisiopatologia e valorização de achados clínicos e de imagem. O Quadro 5.2 mostra as síndromes clínicas mais frequentes na prática do eletrofisiologista e as provas neurofisiológicas mais indicadas para o estudo de cada situação.
1. Barker AT, Jalinous R, Freeston IL. Non-invasivemagnetic stimulation of human motor cortex. Lancet. 1985 May 11;1(8437):1106-7.
2. Brasil-Neto JP, Valls-Solé J, Pascual-Leone A, Cammarota A, Amassian VE, Cracco R, et al. Rapid modulation of human cortical motor outputs following ischaemic nerve block. Brain. 1993 Jun;116 ( Pt 3):511-25.
3. Brown P, Rothwell JC, Thompson PD, Britton TC, Day BL, Marsden CD. New observations on the normal auditory startle reflex in man. Brain. 1991 Aug;114 (Pt 4):1891-902.
4. Buchthal F. An introduction to electromyography. Copenhagen: Scandinavian University; 1957.
5. Chokroverty S. Magnetic stimulation in clinical neurophysiology. Boston: Butterworths; 1989.
6. Cruccu G, Iannetti GD, Marx JJ, Thoemke F, Truini A, Fitzek S, et al. Brainstem reflex circuits revisited. Brain. 2005 Feb;128(Pt 2):386-94.
7. Daube JR. Application of quantitative methods in neuromuscular disorders. In: Halliday AM, Buttler SR, Paul R, editors. A textbook of clinical neurophysiology. New York: Wiley and Sons; 1987.
8. Dawson GD, Scott JW. The recording of nerve action potentials through the skin in man. J Neurol Psychiat.1949;12:259-67.
9. Dumitru D. Electrodiagnostic medicine. Philadelphia: Hanley & Belfus; 1995. Generalized peripheral neuropathies; p.741-850.
10. Kimura J, Powers JM, Van Allen MW. Reflex response of orbicularis oculi muscle to supraorbital nerve stimulation. Study in normal subjects and in peripheral facial paresis. Arch Neurol. 1969 Aug;21(2):193-9.
11. Kimura J. Disorder of interneurons in Parkinsonism. The orbicularis oculi reflex to paired stimuli. Brain.1973;96(1):87-96.
12. Kobayashi M, Pascual-Leone A. Transcranial magnetic stimulation in neurology. Lancet Neurol. 2003Mar;2(3):145-56.
13. Lefaucheur JP, Créange A. Neurophysiological testing correlates with clinical examination according to fibre type involvement and severity in sensory neuropathy. J Neurol Neurosurg Psychiatry. 2004 Mar;75(3):417-22.
14. MacKay WA, Bonnet M. CNV, stretch reflex and reaction time correlates of preparation for movement direction and force. Electroencephalogr Clin Neurophysiol. 1990 Jul;76(1):47-62.
15. Montagna P, Liguori R, Zappia M. Sympathetic skin response. J Neurol Neurosurg Psychiatry. 1985May;48(5):489-90.
16. Navarro X. Evaluación del sistema nervioso autónomo: XLIV Reunión de la Sociedad Española de Neurología. Barcelona: Prous; 1992.
17. Nickerson RS. Intersensory facilitation of reaction time: energy summation or preparation enhancement? Psychol Rev. 1973 Nov;80(6):489-509.
18. Ongerboer de Visser BW, Goor C. Jaw reflexes and masseter electromyograms in mesencephalic and pontine lesions: an electrodiagnostic study. J Neurol Neurosurg Psychiatry. 1976 Jan;39(1):90-2.
19. Ongerboer de Visser BW, Cruccu G, Manfredi M, Koelman JH. Effects of brainstem lesions on the masseter inhibitory reflex. Functional mechanisms of reflex pathways. Brain. 1990 Jun;113 (Pt 3):781-92.
20. Rothwell JC, Thompson PD, Day BL, Boyd S, Marsden CD. Stimulation of the human motor cortex through the scalp. Exp Physiol. 1991 Mar;76(2):159-200.
21. Shahani B. The human blink reflex. J Neurol Neurosurg Psychiatry. 1970 Dec;33(6):792-800.
22. Scherer K, Bedlack RS, Simel DL. Does this patient have myasthenia gravis? JAMA. 2005 Apr 20;293(15):1906-14.
23. Schestatsky P, Valls-Solé J, Costa J, León L, Veciana M, Chaves ML. Skin autonomic reactivity to thermoalgesic stimuli. Clin Auton Res. 2007a Dec;17(6):349-55.
24. Schestatsky P, Kumru H, Valls-Solé J, Valldeoriola F, Marti MJ, Tolosa E, et al. Neurophysiologic study of central pain in patients with Parkinson disease. Neurology. 2007b Dec 4;69(23):2162-9.
25. Schieppati M. The Hoffmann reflex: a means of assessing spinal reflex excitability and its descending control in man. Prog Neurobiol. 1987;28(4):345-76.
26. Stalberg E, Trontelj JV. Single fiber electromyography. Surrey (UK): Mirvalle; 1979.
27. Topka H, Hallett M. Perioral reflexes in orofacial dyskinesia and spasmodic dysphonia. Muscle Nerve. 1992 Sep;15(9):1016-22.
28. Valls-Sole J, Graus F, Font J, Pou A, Tolosa ES. Normal proprioceptive trigeminal afferents in patients with Sjögren’s syndrome and sensory neuronopathy. Ann Neurol. 1990 Dec;28(6):786-90.
29. Valls-Solé J, Lou JS, Hallett M. Brainstem reflexes in patients with olivopontocerebellar atrophy. Muscle Nerve. 1994 Dec;17(12):1439-48.
30. Valls-Sole J, Monforte R, Estruch R. Abnormal sympathetic skin response in alcoholic subjects. J Neurol Sci. 1991 Apr;102(2):233-7.
31. Valls-Solé J, Solé A, Valldeoriola F, Muñoz E, Gonzalez LE, Tolosa ES. Reaction time and acoustic startle in normal human subjects. Neurosci Lett. 1995 Aug 4;195(2):97-100.
32. Veciana M, Valls-Solé J, Schestatsky P, Montero J, Casado V. Abnormal sudomotor skin responses to temperature and pain stimuli in syringomyelia. J Neurol. 2007 May;254(5):638-45.
33. Woodruff-Pak DS, Thompson RF. Classical conditioning of the eyeblink response in the delay paradigm in adults aged 18-83 years. Psychol Aging. 1988 Sep;3(3):219-29.
http://www.abemexam.org/
http://www.ifcn.info/
http://www.sbnc.org.br/
O MedicinaNET é o maior portal médico em português. Reúne recursos indispensáveis e conteúdos de ponta contextualizados à realidade brasileira, sendo a melhor ferramenta de consulta para tomada de decisões rápidas e eficazes.
| Medicinanet Informações de Medicina S/A Cnpj: 11.012.848/0001-57 | info@medicinanet.com.br |
MedicinaNET - Todos os direitos reservados.
Em função da pandemia do Coronavírus informamos que não estaremos prestando atendimento telefônico temporariamente. Permanecemos com suporte aos nossos inscritos através do e-mail info@medicinanet.com.br.